Estructura Tridimensional y Funcional de las Proteínas
Las proteínas son moléculas formadas por aminoácidos que están unidos por un tipo de enlaces conocidos como enlaces peptídicos. El orden y la disposición de los aminoácidos dependen del código genético de cada persona. Una proteína puede ser una sola cadena polipeptídica o puede estar formada por varias de esas cadenas unidas entre sí por interacciones débiles.
Las moléculas individuales de proteína se pueden describir mediante hasta cuatro niveles de estructura. Las secuencias de aminoácidos siempre se escriben desde el amino terminal (N-terminal) hasta el carboxilo terminal C- (C-terminal). La estructura tridimensional de una proteína se describe con tres niveles adicionales: estructura secundaria, estructura terciaria y estructura cuaternaria. Las fuerzas que mantienen, o estabilizan, estos tres niveles son no covalentes, de manera primordial. La estructura secundaria se refiere a las regularidades en las conformaciones locales mantenidas por puentes de hidrógeno entre los hidrógenos de amida y los oxígenos de carbonilo en la columna vertebral del péptido. Las estructuras secundarias principales son las hélices a y las hebras b (incluyendo las láminas a). Se acostumbra representar las regiones helicoidales a con dibujos que muestran las estructuras de proteínas plegadas; las hebras b se representan con flechas anchas que apuntan desde la dirección N-terminal hacia la C-terminal. La estructura terciaria describe la cadena polipeptídica totalmente plegada y compactada. Muchos polipéptidos plegados consisten en varias unidades distintas unidas por un tramo corto de residuos de aminoácidos; a dichas unidades se les conoce como dominios. Las estructuras terciarias se estabilizan por las interacciones de cadenas laterales de aminoácidos en regiones no vecinas de la cadena polipeptídica. La formación de la estructura terciaria acerca partes lejanas de las estructuras primaria y secundaria. Algunas proteínas poseen estructura cuaternaria, que implica la asociación de dos o más cadenas polipeptídica en una multisubunidad, o proteína oligomérica u oligómera. Las cadenas polipeptídicas de una proteína oligómera pueden ser idénticas o distintas.
Las estructuras tridimensionales de biopolímeros como las proteínas pueden determinarse por cristalografía de rayos X y espectroscopia de RMN.
Conformación de enlaces peptidicos
la estructura de las proteínas comienza con la de los enlaces peptídicos, o enlaces de péptido, que unen a los aminoácidos en una cadena polipeptídica. Los dos átomos que intervienen en el enlace peptídico, junto con sus cuatro sustituyentes (el átomo de oxígeno carbonílico, el átomo de hidrógeno de amida y los dos átomos adyacentes de carbono a) constituyen el grupo peptídico. Los análisis cristalográficos de pequeños péptidos con rayos X revelan que el enlace entre el carbono carbonílico y el nitrógeno es más corto que un enlace sencillo típico C—N, pero más largo que los dobles enlaces C= N típicos. Además, el enlace entre el carbono carbonílico y el oxígeno es un poco mayor que el doble enlace típico C =O. Esas mediciones indican que los enlaces peptídicos tienen ciertas propiedades del enlace doble y se pueden representar mejor como un híbrido de resonancia.
La hélice α
La conformación helicoidal a fue propuesta por Linus Pauling y Robert Corey en 1950. Tuvieron en cuenta las dimensiones de los grupos peptídicos, las posibles restricciones estéricas y las oportunidades de estabilización por formación de puentes de hidrógeno. Su modelo explicó la principal repetición observada en la estructura de la c-queratina, una proteína fibrosa. Sucede que esta repetición de 0.50 a 0.55 nm es el paso (la distancia axial por cada vuelta) de la hélice a. Max Perutz agregó otros apoyos a la estructura al observar una unidad repetitiva secundaria de 0.15 nm en la figura de difracción de rayos X de la queratina a. Esta repetición de 0.15 nm corresponde a la elevación o subida de la hélice a (la distancia que cada residuo en la hélice avanza a lo largo de su eje). Perutz también demostró que la hélice a estaba presente en la hemoglobina y con ello confirmó que esta conformación existe en proteínas globulares más complejas.
Hebra ß y Laminas ß
La otra estructura secundaria común se llama estructura ß, una clase que incluye a hebras ß y láminas ß. Las hebras B son partes de la cadena polipeptídica que se encuentran casi totalmente extendidas. Cada residuo en una hebra b ocupa de 0.32 a 0.34 nm de la longitud total, en contraste con la espiral compacta de una hélice a, donde cada residuo corresponde a 0.15 nm de la longitud general. Cuando se ordenan varias hebras ß lado a lado forman láminas ß, estructura que propusieron originalmente Pauling y Corey cuando desarrollaban el modelo teórico de la hélice a. Las proteínas casi nunca contienen hebras b aisladas porque la estructura en sí no es mucho más estable que otras conformaciones. Sin embargo, las láminas ß se hallan estabilizadas por puentes de hidrógeno entre los oxígenos carbonílicos y los hidrógenos de amida en hebras ß adyacentes. Así, en las proteínas, las regiones de estructura ß casi siempre se encuentran en láminas. Las hebras ß con puentes de hidrógeno pueden estar en cadenas separadas de polipéptidos o en diferentes segmentos de la misma cadena. Las hebras ß en una lámina pueden ser paralelas (con la misma dirección de N a C-terminal) o antiparalelas (con direcciones opuestas de N a C-terminal). Cuando las hebras b son antiparalelas, los puentes de hidrógeno son casi perpendiculares a las cadenas extendidas del polipéptido. En el ordenamiento paralelo, los puentes de hidrógeno no son perpendiculares a las cadenas extendidas y cada residuo forma puentes de hidrógeno con los grupos carbonilo y amida de dos residuos diferentes en la cadena adyacente. Las láminas paralelas son menos estables que las antiparalelas, quizá porque los puentes de hidrógeno están distorsionados en el ordenamiento paralelo. A veces, a la lámina ß se le llama lámina B plegada ya que los grupos peptídicos planos se encuentran entre sí formando ángulos como en el plisado de un acordeón. El resultado de los ángulos de enlace entre grupos de péptidos es que las cadenas laterales de aminoácido apuntan arribßa y abajo, alternadamente, del plano de la lámina. Una lámina ß típica contiene desde dos hasta 15 hebras individuales. Cada hebra está constituida por un promedio de seis residuos de aminoácido.
a) Lámina b paralela. Los puentes de hidrógeno están a distancias uniformes, pero están inclinados.
Aminoácidos y La Estructura Primaria de las Proteínas
Los aminoácidos son compuestos orgánicos que se combinan para formar proteínas. Los aminoácidos y las proteínas son los pilares fundamentales de la vida.
Las funciones biológicas de las proteínas son las siguientes:
1. Muchas proteínas actúan como enzimas, los catalizadores bioquímicos. Las enzimas catalizan casi todas las reacciones que suceden en los organismos vivos.
2. Algunas proteínas se unen con otras moléculas para su almacenamiento y transporte. Por ejemplo, la mioglobina se enlaza con el oxígeno en las células de los músculos esquelético y cardiaco, mientras que la hemoglobina se une y transporta al O2 y al CO2 en los glóbulos rojos.
3. Algunas proteínas, como la tubulina, actina y colágena, proporcionan soporte y forma a las células, y en consecuencia a los tejidos y los organismos.
4. Los conjuntos de proteínas pueden efectuar trabajo mecánico, como el movimiento de flagelos, la separación de cromosomas en la mitosis y la contracción de los músculos.
5. Muchas proteínas desempeñan un papel en la descodificación de la información celular. Algunas intervienen en la traducción, mientras que otras juegan un papel en la regulación de la expresión genética al unirse a los ácidos nucleicos.
6. Algunas proteínas son hormonas que regulan las actividades bioquímicas en las células o los tejidos blancos; otras proteínas sirven como receptores de las hormonas.
7. Algunas proteínas desarrollan funciones muy especializadas. Por ejemplo, los anticuerpos defienden a los vertebrados contra infecciones bacterianas y víricas, y las toxinas, que producen algunas bacterias, matan organismos mayores.
Todos los organismos emplean los mismos 20 aminoácidos como bloques constructivos para armar las moléculas de proteína. A estos 20 aminoácidos se les llama aminoácidos comunes, estándar o normales. A pesar de la poca cantidad de los aminoácidos, se puede obtener una variedad enorme de distintos polipéptidos al unir los 20 aminoácidos comunes para formar distintas combinaciones. Los aminoácidos se llaman así porque son derivados aminados de ácidos carboxílicos. En los 20 aminoácidos comunes, los grupos amino y carboxilo están unidos al mismo átomo de carbono: el átomo de carbono a. Así, todos los aminoácidos estándar que contienen las proteínas son a-aminoácidos. Al carbono a se unen otros dos sustituyentes: un átomo de hidrógeno y una cadena lateral (R) que es única para cada aminoácido. En los nombres químicos de los aminoácidos, los átomos de carbono se identifican con números que comienzan en el átomo de carbono del grupo carboxilo. [El nombre químico correcto, o nombre sistemático, se apega a reglas establecidas por la Unión Internacional de Química Pura y Aplicada (IUPAC, de International Union of Pure and Applied Chemistry) y la Unión Internacional de Bioquímica y Biología Molecular (IUBMB, de International Union of Biochemistry and Molecular Biology)
Las estructuras de los 20 aminoácidos que suelen encontrarse en las proteínas se muestran en las figuras siguiente:
A. Grupos Alifáticos
La glicina (Gly, G) es el aminoácido más pequeño porque su grupo R no es más que un átomo de hidrógeno; en consecuencia, el carbono a de la glicina no es quiral. Los dos átomos de hidrógeno del carbono ade la glicina imparten poco carácter hidrofóbico a la molécula. Después habrá oportunidad de comprobar que la glicina desempeña un papel único en la estructura de muchas proteínas porque su cadena lateral es lo bastante pequeña como para tener cabida en nichos en los que a otros aminoácidos les resultaría imposible hacerlo.
Hay cuatro aminoácidos: alanina (Ala, A), valina (Val, V), leucina (Leu, L) y el isómero estructural de la leucina, isoleucina (Ile, I), que tienen cadenas laterales alifáticas saturadas. La cadena lateral de la alanina es un grupo metilo, mientras que la valina presenta una cadena lateral ramificada con tres carbonos, y la leucina y la isoleucina contienen una cadena lateral ramificada de cuatro carbonos cada una. Los átomos de carbono a y b de la isoleucina son asimétricos. Ya que la isoleucina tiene dos centros quirales, posee cuatro estereoisómeros posibles.
B. Grupos Aromáticos
La fenilalanina (Phe, F), tirosina (Tyr, Y) y el triptófano (Trp, W) presentan cadenas laterales con grupos aromáticos. En el caso de la fenilalanina es una cadena hidrofóbica bencílica. La tirosina se parece estructuralmente a la fenilalanina; en la tirosina, un grupo hidroxilo sustituye al hidrógeno para de la fenilalanina lo que la convierte en un fenol. El grupo hidroxilo de la tirosina es ionizable, pero bajo condiciones fisiológicas normales retiene su hidrógeno. La cadena lateral del triptófano contiene un grupo indol bicíclico. La tirosina y el triptófano no son tan hidrofóbicos como la fenilalanina porque en sus cadenas laterales hay grupos polares. Los tres aminoácidos aromáticos absorben luz ultravioleta (UV) porque, a diferencia de los aminoácidos alifáticos, los aromáticos contienen electrones p deslocalizados. A pH neutro tanto el triptófano como la tirosina absorben luz a una longitud de onda de 280 nm, mientras que la fenilalanina es casi transparente a 280 nm y absorbe débilmente luz a 260 nm. Dado que la mayor parte de las proteínas contienen triptófano y tirosina éstas absorben luz a 280 nm. La absorbencia a 280 nm se usa en forma rutinaria para estimar la concentración de proteínas en las soluciones.
C. Grupos Azufrados
La metionina (Met, M) y la cisteína (Cys, C) son los dos aminoácidos azufrados. La metionina contiene un grupo tioéter metilo, no polar, en su cadena lateral, lo que la convierte en uno de los aminoácidos más hidrofóbicos. La metionina desempeña un papel especial en la síntesis de proteínas porque casi siempre representa el primer aminoácido en una cadena de polipéptido. La estructura de la cisteína se parece a la de la alanina, con un átomo de hidrógeno reemplazado por un grupo sulfhidrilo(-SH). Aunque la cadena lateral de la cisteína es algo hidrofóbica, también es muy reactiva. Debido a que el átomo de azufre es polarizable, el grupo sulfhidrilo de la cisteína puede formar puentes de hidrógeno débiles con el oxígeno y el nitrógeno. Además, el grupo sulfhidrilo de la cisteína es un ácido débil, lo cual le permite perder un protón y transformarse en un ion tiolato con carga negativa.
D. Cadenas Laterales con grupo Alcohol
La serina (Ser, S) y la treonina (Thr, T) tienen cadenas laterales polares sin carga que contienen grupos b-hidroxilo. Estos grupos alcohol dan carácter hidrofílico a las cadenas laterales alifáticas. Además de la cadena lateral fenólica más ácida de la tirosina, los grupos hidroxilo de la serina y la treonina despliegan las propiedades débiles de ionización de los alcoholes primarios y secundarios. El grupo hidroximetilo de la serina (—CH2OH) no se ioniza en forma apreciable en soluciones acuosas; empero, este alcohol puede reaccionar dentro de los sitios activos de varias enzimas como si estuviera ionizado. La treonina, como la isoleucina, cuenta con dos centros quirales, los átomos de carbono a y b. La L-treonina es el único de los cuatro estereoisómeros que se encuentra con frecuencia en las proteínas. (Los otros estereoisómeros son D-treonina, L-alotreonina y D-alotreonina).
E. Grupo R Basico
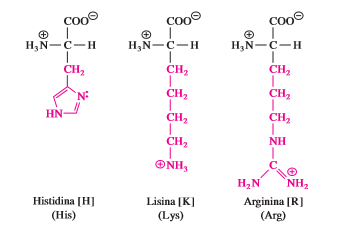
La histidina (His, H), lisina (Lys, K) y arginina (Arg, R) presentan cadenas laterales hidrofílicas que son bases nitrogenadas y tienen carga positiva a pH 7. La cadena lateral de la histidina contiene un sustituyente de anillo de imidazol. La forma protonada de este anillo se llama ion imidazolio (sección 3.4). La lisina es un diaminoácido y tiene grupos amino a y e al mismo tiempo. El grupo e-amino existe como ion alquilamonio (—CH2 —NH3 ) a pH neutro y confiere una carga positiva a las proteínas. La arginina es el más básico de los 20 aminoácidos porque su cadena lateral de ion guanidinio está protonada bajo todas las condiciones que se encuentran de manera habitual dentro de una célula. Las cadenas laterales de arginina también aportan cargas positivas a las proteínas.
F. Grupos R acidos y sus amidas derivadas
El aspartato (Asp, D) y el glutamato (Glu, E) son aminoácidos dicarboxílicos y tienen cadenas laterales hidrofílicas con carga negativa a pH 7. Además de los grupos carboxilo a, el aspartato posee un grupo carboxilo b y el glutamato un grupo carboxilo g. El aspartato y el glutamato confieren carga negativa a las proteínas porque sus cadenas laterales se encuentran ionizadas a pH 7. A veces se les llama ácido aspártico y ácido glutámico, respectivamente, pero bajo la mayoría de las condiciones fisiológicas se encuentran como bases conjugadas y, al igual que otros carboxilatos, tienen el sufijo ato. Es probable que el glutamato sea conocido bajo la forma de su sal monosódica, el glutamato monosódico (MSG, de monosodium glutamate), que se usa en alimentos como intensificador de sabores. La asparagina (Asn, N) y la glutamina (Gln, Q) son las amidas del ácido aspártico y el ácido glutámico, respectivamente. Aunque las cadenas laterales de la asparagina y la glutamina son eléctricamente neutras, estos aminoácidos son muy polares y se encuentran con frecuencia en las superficies de las proteínas, donde pueden interactuar con moléculas de agua. Los grupos amida polares de la asparagina y la glutamina también pueden formar puentes de hidrógeno con las cadenas laterales de otros aminoácidos polares.
II. Unión de los aminoácidos por enlaces peptidicos en las proteínas
La secuencia lineal de aminoácidos en una cadena polipeptídica se llama estructura primaria de una proteína. A los niveles más altos de estructura se les llaman estructura secundaria, terciaria y cuaternaria. La estructura de las proteínas se describirá con más detalle en el siguiente capítulo, pero es importante comprender los enlaces peptídicos y la estructura primaria antes de describir algunos de los temas restantes en este capítulo. El enlace que se forma entre los aminoácidos es un enlace de amida y se llama enlace peptídico, o enlace de péptido. Esta unión se puede concebir como el resultado de una condensación simple del grupo carboxilo a de un aminoácido con el grupo amino a del otro. Observe que se pierde una molécula de agua de los aminoácidos que se condensan en la reacción.. A diferencia de los grupos carboxilo y amino de los aminoácidos libres en solución, los grupos que intervienen en los enlaces peptídicos no tienen cargas iónicas. Las mitades de aminoácido unidas en una cadena polipeptídica se llaman residuos de aminoácido. Los nombres de los residuos se forman sustituyendo la terminación -ina o -ato por -ilo (o -il, en nombres compuestos). Por ejemplo, un residuo de glicina en un polipéptido se llama glicilo y un residuo de glutamato se llama glutamilo. En los casos de asparagina, glutamina y cisteína, -ilo sustituye la –a final para formar asparaginilo, glutaminilo y cisteinilo, respectivamente. La terminación ilo indica que el residuo es una unidad de acilo (estructura que carece del hidroxilo del grupo carboxilo). El grupo amino libre y el grupo carboxilo libre en los extremos opuestos de una cadena de péptido se llaman N-terminal (o terminal N, terminal amino) y C-terminal (o terminal C, terminal carboxilo), respectivamente. Por convención, los residuos de aminoácido en una cadena peptídica se numeran desde el N-terminal hasta el C-terminal y se suelen escribir de izquierda a derecha. Esta convención corresponde a la dirección de la síntesis de la proteína. La síntesis comienza con el aminoácido N-terminal —que casi siempre es metionina— y prosigue en secuencia hacia el C-terminal, agregando uno por uno los residuos.
El
agua es la molécula más importante para la vida, ya que constituye alrededor
del 60 -90% del peso de las células. En
el ámbito bioquímico es importante comprender al agua y sus propiedades, porque
mediante ella podemos determinar cómo las macromoléculas asumensus formas características como respuesta al
agua. Algunos tipos de moléculas interaccionan en forma extensa con el agua y
en consecuencia son muy solubles.
El
agua es una molécula formada por dos átomos de oxígeno y un átomo de hidrogeno
(H2O)
formado por un Angulo de 104.5º entre
dos enlaces covalentes. El núcleo de oxígeno de esta molécula atrae a los
electrones más que el protón único del núcleo del hidrógeno, por ende, los átomos de oxígeno son más electronegativos
que los de hidrógeno. El resultado es que se suscita una distribución desigual
de cargas dentro de cada enlace O—H en la molécula de agua, con el oxígeno
teniendo una carga negativa parcial (-) y el hidrógeno con una carga positiva
parcial (+). Esta distribución desigual de la carga dentro del enlace se llama
dipolo y se dice que el enlace es polar. Una de las consecuencias importantes
de la polaridad de la molécula de agua es que dichas moléculas se atraen entre
sí.
Una
de las excelentes propiedades físicas que tiene el agua es que actúa como
disolvente de la mayoría de los compuestos, ya que posee una viscosidad
intrínseca que no impide mucho el movimiento de las moléculas disueltas en
ella. Esta puede interactuar y disolver otros compuestos polares y compuestos
que se ionizan. La ionización se relaciona con la ganancia o pérdida de un
electrón, que da lugar a un átomo o a un compuesto que presenta una carga neta.
Las moléculas que se pueden disociar y formar iones se llaman electrólitos. En
base a la solubilidad del agua se puede deducir lo siguiente:
-Las sustancias
que se disuelven con facilidad en el agua se dice que son a fines de agua (hidrofilicas).
- Las sustancias
que no son solubles en agua se llaman hidrófobas.
-Las sustancias
que tienen tanto una parte soluble como
una insoluble, como en el caso de los fosfolípidos, se dice que son
antipáticas.
Para
las moléculas pequeñas, la velocidad de difusión dentro de las células es más o
menos la cuarta parte de la que muestran en agua pura. Para las moléculas
grandes, como las de proteínas, la tasa de difusión en el citoplasma disminuye
hasta a 5-10% de la velocidad en agua. Esta desaceleración se debe en gran
parte al hacinamiento molecular. Esta molécula es importantes para procesos
celulares como:
La bioquímica es el estudio de las moléculas y las reacciones químicas de la vida. Es la disciplina que emplea los principios y el lenguaje de la química a fin de explicar la biología a nivel molecular. Los bioquímicos descubrieron que los compuestos químicos y los procesos metabólicos centrales son los mismos que se encuentran en organismos tan distantes como las bacterias, plantas y humanos. Se sabe que los principios básicos de la bioquímica son comunes a todos los organismos vivos. Aunque en la práctica los científicos concentran sus esfuerzos de investigación en organismos particulares, sus resultados se pueden extrapolar a muchas otras especies.
La bioquímica surgió como ciencia dinámica tan sólo desde hace 100 años. No obstante, las bases para el campo de trabajo que dieron pie al surgimiento de la bioquímica como ciencia moderna fueron sentadas desde hace muchos siglos. El periodo anterior al siglo XX fue testigo de rápidos avances en la comprensión de los principios químicos básicos como la cinética de reacción y la composición atómica de las moléculas. Para fines del siglo XIX se habían identificado numerosas sustancias químicas producidas por los organismos vivos. Desde entonces, la bioquímica se ha convertido en una disciplina organizada y los bioquímicos dilucidaron muchos de los procesos químicos de la vida. El crecimiento de la bioquímica y su influencia en otras disciplinas seguirá su marcha durante el siglo XXI.
En bioquímica también son importantes las macromoleculas biológicas, las cuales forman un polímero creado mediante la unión de muchas moléculas orgánicas más pequeñas, o monómeros, por medio de condensaciones (la remoción de los elementos del agua). Cada monómero incorporado a una cadena macromolecular se denomina residuo. Acontinuacion se presentara una reseña breve,